Оргинальная статья
РАСТИТЕЛЬНОСТЬ И РАСТИТЕЛЬНОЕ ВЕЩЕСТВО ПЛОСКОБУГРИСТЫХ ТОРФЯНИКОВ
Н.П. Косых1  , Н.П.Миронычева-Токарева1, Е.В.Михайлова1, 2, Л.Г.Колесниченко3
, Н.П.Миронычева-Токарева1, Е.В.Михайлова1, 2, Л.Г.Колесниченко3
Адрес: 1ФГБУН Институт почвоведения и агрохимии СО РАН, проспект Академика Лаврентьева, 8/2, г. Новосибирск, 630090, Россия
2 Сибирский государственный университет геосистем и технологий, ул. Плахотного 10, г. Новосибирск, 630108, Россия
3Томский государственный университет, Научно исследовательский институт биологии и биофизики, ул. Ленина 25, г. Томск, 634050, Россия
Цель исследования. Выявить особенности распределения запасов растительного вещества и продукции в зависимости от состава растительного покрова, характеристики почвенного слоя и микрорельефа плоскобугристых болот северной тайги Западной Сибири.
Место и время проведения. Плоскобугристые мочажинно-озерковые комплексы на мерзлых торфяниках подзоны северной тайги Западной Сибири. Приведены результаты многолетних исследований с 2013 по 2017 гг.
Методология. Комплексные исследования почвенного и растительного покрова. Изучена структура растительного вещества с выделением живых и мертвых подземных органов и чистая первичная продукция болотных экосистем.
Основные результаты. Чистая первичная продукция экосистем плоскобугристого болота колебалась в пределах от 3,0 до 4,5 т/га в год в зависимости от видового состава растительного сообщества. В среднем составила около 4,0 т/га в год при запасе живой фитомассы – 14,8 т/га, в спущенных озерах – 8,5 т/га в год, при запасах фитомассы – 18,0 т/га. На плоских буграх отношение надземной продукции кустарничков к подземной 1:4, в мочажинах преобладали подземные органы осок и пушиц 1:6. Максимальный вклад подземной продукции наблюдалсяя в мезотрофных мочажинах и составлял 70% от общей продукции и создавался, в основном, подземными органами крупных осок и пушиц. Общая первичная продукция на буграх составляла 4,1 т/га в год, в олиготрофных мочажинах – 3,0 т/га в год, в хасыреях достигала максимальной величины для этой подзоны – 8,5 т/га в год.
Заключение. Анализ полученного материала показал, что средние значения запасов растительного вещества как в надземной, так и в подземной сфере были в течении ряда лет на одном уровне и близки запасам полученным для болот Канады, Швеции и Финляндии. Распределение запасов фитомассы и годичной продукции в плоскобугристых болотах северной тайги зависили от условий минерального питания почвы и обводненности корнеобитаемого слоя. Доминирующая роль мхов в продукции экосистемы сохранялась только в олиготрофных мочажинах, на буграх преобладали лишайники, видовое разнообразие которых превосходит остальные группы растений. При равных условиях в мочажинах запасы фитомассы и годичная продукция растут с увеличением трофности почвы, а в экосистемах бугров уменьшаются при наличии мерзлого слоя почвы. Анализ продуктивности на разных болотных почвах северной тайги позволяет сделать вывод, что почвенные условия оказывают существенное влияние на интенсивность продукционного процесса и обусловливают в целом невысокий уровень продуктивности. Запасы и продукция фитомассы на буграх лимитированы небольшой глубиной протаивания, в летний сезон сюда добавляется иссушение верхнего слоя, его прогревания, что при близком залегании мерзлоты создает неблагоприятные условия для жизни корневых систем.
Ключевые слова:
торфяники; мерзлота; биологическая продуктивность; растительное вещество; запасы; фитомасса; мортмасса; чистая первичная продукцияЦитирование: Косых Н.П., Миронычева-Токарева Н.П., Михайлова Е.В., Колесниченко Л.Г. Растительность и растительное вещество плоскобугристых торфяников // Почвы и окружающая среда. 2019. Том 2. № 1. e55. doi: 10.31251/pos.v2i1.55
Введение
С экологической точки зрения любое преобразование поверхности земли сопровождается трансформацией почвенно-растительного покрова с формированием новых экосистем, состояние которых определяется формой поверхности и составом ее почвенно-растительного комплекса. Любая экосистема характеризуется структурой с набором компонентов со связями между ними и их количественными характеристиками на определенный момент времени или в среднем за период. Одной из особенностей болотных экосистем является их ранимость при антропогенных воздействиях, приводящих к их деградации (Тыртиков, 1972; Тишков, 1986; Москаленко, 1999). Чтобы предотвратить или уменьшить негативное влияние на компоненты экосистемы, необходимо при планировании работ по природопользованию рассматривать характеристики компонентов с точки зрения совокупности состава микрокомплексов в экосистеме, учитывая характер их взаимосвязи между собой и воздействие абиотических факторов среды для последующего прогнозирования процессов восстановления почвенного и растительного покрова. В настоящее время оценены запасы фитомассы и ее ежегодное воспроизводство в наземных природных экосистемах, слагающих растительный покров административных регионов Западной Сибири. Несмотря на то, что универсальность системы обменных процессов биологического круговорота, включающей в себя определенные запасы вещества и интенсивности процессов, связывающих компоненты круговорота, установлена, степень сходства будет различна в разных типах экосистем (Базилевич, Титлянова, 2008).
Болота криолитозоны Западной Сибири представлены комплексами плоских бугров с олиготрофными бедными мочажинами и заторфованными котловинами спущенных термокарстовых озер (хасыреев) с сочетанием травяно-сфагново-кустарничковых экосистем, мелких озерков и формирующихся бугров пучения (Брыксина, Полищук, 2015). Установлено, что на границе перехода от экосистем бугров к мочажинам и спущенным озерам (хасыреям) количественные характеристики запасов фракций фитомассы определяются типом почвы и видовым составом растительного сообщества, которые зависят от положения в мезо- и микрорельефе ландшафта. Для моделирования процессов функционирования заболоченных ландшафтов Западной Сибири при разных климатических сценариях в болотных экосистемах бугров, олиготрофных мочажин и евтрофных хасыреев во времени требуется продолжение работ по определению составляющих биотического круговорота. Цель данной работы состояла в выявлении особенностей распределения запасов растительного вещества и продукции в зависимости от состава растительного и почвенного покровов, а также микрорельефа плоскобугристых болот северной тайги Западной Сибири.
Материалы и методы
Ключевой участок, репрезентативный для территории Сибирских Увалов, находится в подзоне северной тайги Западной Сибири и относится к Обско-Тазовскому региону мерзлых комплексных торфяных болот (Мельцер, 1985). Рельеф полого-гривисто-увалистый. Около 50% территории заболочено. Болотные массивы заняты полого-бугристыми мочажинно-озерковыми болотными комплексами на мерзлых торфяниках. Вдоль мелких болотных речек развиты крупнобугристые болота. Благодаря высокой заболоченности и небольшой локальной амплитуде высот рельефа, грунтовые воды держатся в пределах верхних 5-10 метров, а на периферии плоских повышений - около 2-3 метров. Удерживаемая торфяниками болот вода позволяет поддерживать уровень грунтовых вод на достаточно высоком уровне. Микрорельеф болот определен мерзлотными процессами. Бугры местами достигают 4-6 м, но в основном их высота не превышает 1-2 м. Гривы представляют собой плосковершинные холмы и гряды 1-2 м высотой. Они сложены суглинистыми грунтами озерного происхождения. Часть территории покрыта супесями и опесчаненными суглинками, часто имеющими примесь гальки. Микропонижения заболочены, и частично заняты озерками, соединенными мелкими протоками в цепочки и создающие системы плоскобугристых болот. Большое влияние на развитие мезо-, микро- и нано- рельефа оказывает многолетняя мерзлота. С ней связаны криогенные процессы, создающие в зоне северной тайги своеобразные формы рельефа и обуславливающие сложные комплексы почвенного покрова (Смоленцев, 2002).
Климат территории резко континентальный, суровый. Средняя годовая температура воздуха -5оС. Годовое количество осадков составляет 580 мм, основная часть которых приходится на теплое время года (с апреля по октябрь). Зима холодная, температура воздуха в январе составляет -20о. Лето сравнительно теплое, однако средняя температура воздуха в июне может колебаться от 4 до 20оС. Первая половина лета сухая, вторая избыточно влажная. Осень холодная, в сентябре температура воздуха переменчива в течении суток (от -5 до 10оС). В октябре температура воздуха становится отрицательной (Павлов, 2008).
Определение запасов и структуры растительного вещества проводили на ключевом участке, расположенном на плоской междуречной равнине, занятой плоскобугристым мерзлым торфяником с комплексом хасыреев, мочажин и озерков. Длина склона изучаемого участка около 140 м, уклоны его не превышают 0,02.
На супесчаных отложениях, перекрытых торфяниками, формируются торфяные олиготрофные почвы. В настоящее время рост торфяных отложений осуществляется в соответствии с условиями микрорельефа, выраженность которого зависит как от исходного рельефа территории, так и развития термокарстовых процессов. В автоморфной позиции процесс торфонакопления лимитирован небольшой глубиной протаивания, в летний сезон сюда добавляется иссушение верхнего слоя, его прогревание, что при близком залегании мерзлоты создает неблагоприятные условия для жизни корневых систем. На мерзлых торфяниках формируются торфяные олиготрофные мерзлотные почвы.
Поверхность плоских бугров волнистая, с большим количеством микропонижений. Высота кочек от 10 до 15 см. Глубина талого слоя на буграх в летнее время залегает на глубине 50 см ниже поверхности лишайников (Рис 1). Растительность представлена на буграх кустарничково-лишайниковым сообществом, в мочажинах – кустарничково-осоково- и пушицево-сфагновыми сообществами (табл. 1).
Таблица 1. Характеристика ключевого участка «Ханымей»
| Экосистема | Растительное сообщество | Координаты | |
|---|---|---|---|
| Плоский бугор | Кустарничково-лишайниковое | N 63°47'29.052" | E 75°33'49.104" |
| Мочажина | Кустарничково-осоково-сфагновое, пушицево-сфагновое | N 63°47'29.826" | E 75°33'44.754" |
| Хасырей | Осоково-сфагновое | N 63°47'27.432" | E 75°33'48.774" |
Результаты исследования
Кустарничковый ярус расположен на микроповышениях плоских бугров и редко поднимается выше 15 см от лишайникового покрова. Доминантом кустарничкового яруса является Ledum decumbens (Auton) Lodd. ex Steud (проективное покрытие около 40%); содоминанты – Betula nana L. и Vaccinium vitis-idaea L., Vaccinium uliginosum L., занимающие по 10% проективного покрытия. Общее проективное покрытие (ОПП) кустарничков составляет 60%. Так же единично встречаются Andromeda polifolia L. и Empetrum nigrum L. Травяной ярус образован Rubus chamaemorus L. (5% ОПП). В мохово-лишайниковом ярусе лишайники занимают господствующее положение и по видовому составу, на них приходится 95% проективного покрытия. Доминантами здесь являются Cladonia stellaris (Opiz) Pouzar et Vězda и C. stygia (Fr.) Ruoss. Встречаются также виды Cetraria nivalis (L.) Ach. , C. cuculata (Bellardi)Ach., C. islandica (L.) Ach., Cladonia rangiferina (L.) Web., C. amaurocrea (Flörke) Schaer., Allectoria ohroleuca (Hoffm) Massal. На сфагновые мхи приходится только 5% ОПП; встречается Sphagnum fuscum (Schimp.) Klinggr и S. capillifolium (Ehrh.) Hedw .
Почвенный разрез заложен на плоском бугре в кустарничково-лишайниковом растительном сообществе. Почва – торфяная олиготрофная мерзлотная (Glacic-Fibric Histosols (Dystric) (Рис. 1)

|

|
Рисунок 1. Растительный покров плоских бугров и мерзлотная торфяная почва на бугре
Описание разреза:
Очес (0 – 7) Цвет зеленовато-светло-серый. Представляет собой отмершие части лишайников. Сухой. Рыхлый. С нижележащим горизонтом связан непрочно. Граница ровная. Переход ясный.
Т1 (7 – 10) Мокрый рыжевато-охристый горизонт, сложенный мохово-кустарничковым торфом, со степенью разложения 50%. Густо пронизан корнями растительности и погребенными стволиками кустарничков, с поверхности отмечаются минеральные примеси. Рыхлый. Свежий. Переход в соседний горизонт резкий по окраске. Граница ровная.
Т2 (10 – 25) Рыжевато-бурый торфяной горизонт. На глубине 12 – 20 см фрагментарные прослойки коричневато-бурого торфа. Минеральные примеси отсутствуют. Рыхлый. Свежий. Переход в соседний горизонт резкий по окраске. Граница ровная.
Т3 (25 – 50) Коричневато-темно-бурый слоистый (буровато-красноватые тонкие прослойки). Имеется незначительная примесь минеральных частиц. Уплотнен. Свежий. Мерзлота с 50 см.
Растительные сообщества мочажин сменяют друг друга в зависимости от уровня обводненности (рис. 2). Пушицево-сфагновое сообщество занимает местообитания с наиболее низким уровнем болотных вод (около 15 см). Обычно такие сообщества окаймляют края больших мочажин или занимают микропонижения между буграми. Травяной ярус представлен в основном Eriophorum russeolum Fries с проективным покрытием до 10%, присутствует также Carex rotundata Wahlend. Часто рядом с пушицей единично встречается Andromeda polifolia. В моховом покрове доминирует Sphagnum balticum (Russ) Russ.ex C.Jens. При продвижении в сторону центра округлой мочажины наблюдается повышение уровня болотных вод, что вызывает исчезновение кустарничков, повышение проективного покрытия C. rotundata в травяном ярусе, которая из субдоминантов переходит в доминанты (5-7% проективного покрытия). В моховом покрове также происходит смена доминантов – преобладающими видами становятся Sphagnum lindbergii Schimp. ex Lindb. и Warnstorfia fluitans (Hedw.) Loeske. Осоково-сфагновые сообщества могут полностью занимать центральные части мочажин или же только окаймлять полосой открытую водную поверхность, находящуюся в самом центре мочажины (Косых и др., 2010).

Рисунок 2. Осоково-сфагновая мочажина
Почвенный разрез заложен в осоково-сфагновой олиготрофной мочажине (Таблица 1).
Почва – торфяная олиготрофная (Fibric Histosols (Dystric)) (Рис. 2).
Описание разреза:
Очес (0 – 20) Остатки кустарничков, осок, пушиц и сфагновых мхов.
Т1 (20 – 35) Мокрый коричнево-бурый горизонтнизкой степени разложенности (5-10 %), с включением полуразложившихся кустарничков и пушиц, отмечаются минеральные примеси. Переход постепенный по окраске.
Т2 (35 – 40) Мокрый, бурый, торфяной слой низкой степени разложенности(5-10 %), количество минеральной примеси по сравнению с предыдущим горизонтом заметно уменьшилось.
Т3 (40-50) Темно-бурый низкой степени разложенности (5-10 %), слегка мажется, отмечается значительное количество минеральных примесей.
Растительный покров хасыреев представлен пушицево-осоково-сфагновыми, разнотравно-осоково-сфагновыми сообществами. Доминируют в сообществе растения травяного яруса Carex rostrata Stokes (10%), C. lasiocarpa Ehrh., Eriophorum polystachion L. (5%), E. vaginatum (5%), E. russeolum Fries (5%), единично встречаются Comarum palustre L. и Menyanthes trifoliata L., доля их в проективном покрытии не превышает 3%. Кустарничковый ярус представлен андромедой Andromeda polifolia (10%) и березкой Betnula nana L. В моховом ярусе доминируют с проективным покрытием по 40% Sphagnum lindbergii S. balticum, единично встречаются S. compactum DC. in Lam. et DC. и S.squarrosum Crome.
Почвенный разрез заложен в осоково-сфагновом растительном сообществе хасырея. (Табл.1). Почва – торфяная евтрофная мерзлотная (Hemic Histosols) (Рис.3).
Описание разреза:
Т1 (0–20) Влажный. Серовато-темно-бурый торфяной горизонт высокой степени разложения (70-80%). Переплетен корнями осок и пушиц. Уплотнен. Граница неясно выраженная, переход постепенный по окраске.
Т2 (20–55) Сырой. Темно-бурый торфяной горизонт высокой степени разложенности (70-80%). Очень плотный, сильно мажется.

Рисунок 3. Торфяная евтрофная мерзлотная почва
Определение запасов фракций растительного вещества в экосистемах проводили стандартными методами с небольшими модификациями.
Во всех экосистемах была изучена структура растительного вещества с выделением живых и мертвых подземных органов. Для этого закладывали десять квадратов (50х50 см) случайным образом. На всех десяти квадратах надземную фитомассу срезали на уровне поверхности лишайников или головок мха и с этих же площадок собирали отмершие листья трав и кустарничков. Ветошь отделяли от зеленой фитомассы, которую разбирали по видам и по фракциям приростов текущего и прошлого годов, а так же на многолетние побеги. Всю надземную и подземную фитомассу высушивали при температуре 60ºС и взвешивали. Для определения запасов мохово-лишайникового яруса, общей подземной растительной массы и подземной массы отдельных видов растений на этих же участках отбирали монолиты 10х10х10 см до глубины 30 см в восьмикратной повторности.
Чистая первичная продукция (NPP) складывалась из надземной продукции трав, кустарничков и мхов (ANP) и продукции подземных органов (BNP). Надземная продукция трав соответствует фракции фотосинтезирующей фитомассы. Надземная продукция кустарничков состоит из фитомассы прироста побегов текущего года с растущими на них листьями (Андреяшкина и др., 1981). Продукцию сфагновых мхов определяли по методике «индивидуальных меток» (Косых и др., 2017), а продукцию лишайников - по изменению морфологических признаков стеблей и веточек (Андреев, 1954). Подземную продукцию трав и кустарничков определяли по приросту корней, корневищ и узлов кущения текущего года (Kosykh et al., 2008).
Степень разложения торфяных слоев определялась микроскопическим методом (ГОСТ 10650-2013). Названия почв приведены в соответствии с классификацией почв России (Классификация …, 2004) и Всемирной базой почвенных ресурсов (IUSS Working Group WRB, 2006).
Результаты исследования
Как показывает морфологическое описание почвенных разрезов, профиль состоит из слоев торфа с различной степенью разложенности. Средняя степень разложения торфяных слоев отмечена для почв плоских бугров. Наиболее высокой степенью разложения характеризуются почвенные горизонты евтрофной почвы хасырея – до 80%. В олиготрофных мочажинах степень разложения колеблется в пределах 5-10%. Мерзлотный горизонт находится на глубине 50 см. Мерзлота льдистая, надмерзлотная часть очень влажная. Следы морозной деформации отсутствуют. Реакция среды в органогенной толще меняется постепенно – от слабокислой к сильнокислой (pH водн. 3,4-4,4). Зольность торфа низкая в слоях Т1-Т3 мерзлотной почвы плоского бугра (разрез1), торфяные прослои, состоят из остатков сфагновых мхов. У почв олиготрофных мочажин показатели зольности высоки (28,7-81,6%) за счет большого количества минеральных примесей. Зольность почв хасырея выше, чем почв плоского бугра и значительно колеблется в пределах от 7,8% до 24,4 %. Торф беден подвижными окислами зольных элементов. Содержание обменных катионов Ca2+ и Mn2+ распределено по профилю всех изучаемых почв неравномерно. Наибольшее количество катионов Ca2+ и Mn2+ отмечается в горизонте Т1 (плоский бугор - 13,3; мочажина - 14,3; хасырей - 20,8). Средние величины содержания катионов в нижележащих слоях изменяются в пределах 7,3-9,0. Характер их распределения говорит об их нисходящем перемещении в периодически замерзающем и оттаивающем профиле почвы (Таблица 2). Анализ морфологических особенностей изучаемых почв показывает, что профили практически идентичны, а основное различие заключается в большей или меньшей степени разложения торфяных горизонтов. По мере увеличения степени разложенности торфа снижается величина кислотности и возрастает степень насыщенности основаниями (Васильевская и др., 1986).
Таблица 2. Свойства почв ключевого участка
|
Горизонт |
Глубина, см |
pH водный |
Зольность, % |
Ca2+ водн. |
Mg2+ водн. |
Ca2+ обмен. |
Mg2+ обмен |
|---|---|---|---|---|---|---|---|
|
мг-экв на 100 г почвы |
|||||||
| Торфяная олиготрофная мерзлотная. Разрез 1 | |||||||
|
Очес |
0 – 6 |
4,4 |
– |
6,7 |
2,4 |
4,7 |
2,1 |
|
Ат |
6 – 10 |
3,7 |
19,1 |
6,6 |
3,8 |
7,8 |
5,2 |
|
Т1 |
10 – 20 |
3,4 |
0,6 |
4,1 |
3,1 |
13,3 |
5,8 |
|
Т2 |
20 – 30 |
3,5 |
0,6 |
6,6 |
3,1 |
9,0 |
2,2 |
|
Т3 |
40 – 50 |
3,4 |
1,7 |
7,5 |
3,2 |
11,4 |
4,2 |
| Торфяная олиготрофная. Разрез 2 | |||||||
|
Очес |
0 – 10 |
4,7 |
28,7 |
10,7 |
3,6 |
7,3 |
4,9 |
|
Т1 |
10 – 20 |
4,1 |
43,5 |
8,0 |
4,0 |
14,3 |
7,7 |
|
Т2 |
20 – 30 |
4,6 |
81,6 |
5,2 |
2,9 |
4,5 |
0,8 |
| Торфяная евтрофная мерзлотная. Разрез 3 | |||||||
|
Очес |
0 – 10 |
4,2 |
9,7 |
10,1 |
5,0 |
13,9 |
6,1 |
|
Т1 |
10 – 20 |
4,3 |
24,4 |
2,9 |
1,0 |
20,8 |
1,6 |
|
Т2 |
20 – 30 |
4,5 |
11,3 |
3,1 |
1,0 |
9,7 |
– |
|
Т2 |
30 – 40 |
4,6 |
7,8 |
4,1 |
1,0 |
9,0 |
2,4 |
|
Т3 |
40–50 |
4,7 |
– |
5,5 |
1,8 |
11,4 |
1,0 |
В торфяной почве бугров на глубине до 6 см запасы корней кустарничков сформировали около половины всех запасов растительного вещества 46% (7 т/га). С глубиной запасы корней снизились до 27% и составили 4,0 т/га. В верхнем слое почвы олиготрофной мочажины (сфагновый очес) подземные органы осок и пушиц составили 2,7 т/га или 34%, на глубине 10-20 (Т1) отмечалось наибольшее количество корней и корневищ: они составили 3,6 т/га, ниже, т.е. в слое Т2, фитомасса корней уменьшилась и составила около1,7 т/га. В евтрофной торфяной почве хасыреев подземные органы осок и пушиц достигали наибольших величин в слое Т1 на глубине 10-20 см - 10 т/га (70% от всех запасов подземных органов), ниже на глубине 20-30 в слое Т2 запасы снижались (Рис. 4).
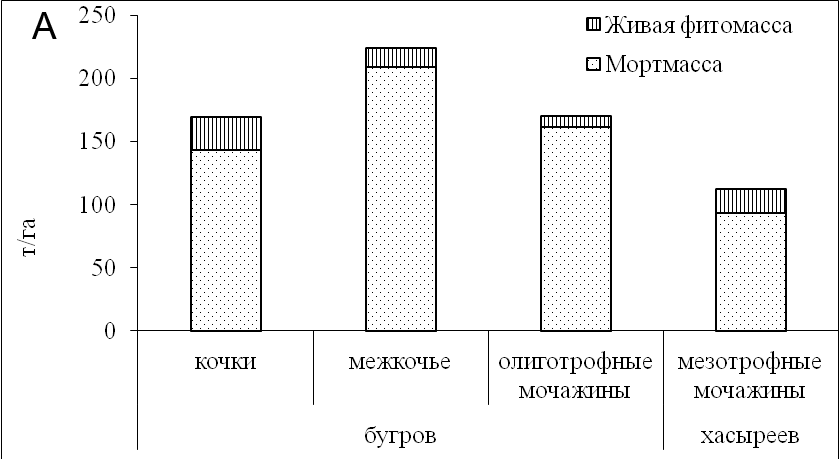
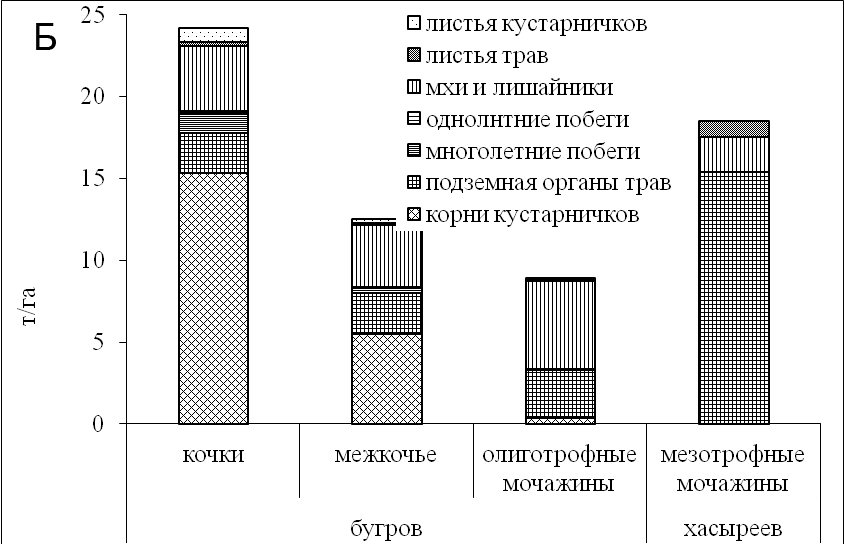
Рисунок 4. Запасы фитомассы и мортмассы (а), структура запасов фитомассы (б) на разных элементах рельефа плоскобугристого болота
Количество мертвого растительного вещества (мортмассы) в среднем составило 180 т/га, накапливаясь в межкочечных пространствах плоскобугристого болота до 210 т/га и уменьшаясь на кочках бугров до 140 т/га. В мочажинах его количество колебалось в пределах от 90 до 160 т/га (рис. 4а). В контрастных, но стабильных системах – кочках бугров и олиготрофных мочажинах - наблюдались меньшие колебания в запасах, чем в межкочьях. Вероятно, это связано с тем, что туда не проникает холодный ветер, что способствует образованию микроклиматических оазисов, вызывающих протаивание мерзлоты на большую глубину, нежели на буграх.
Живое растительное вещество (фитомасса) исследуемых экосистем - это наиболее лабильная фракция, так как зависит от погодных условий текущего года. Минимальные запасы живой фитомассы были в экосистемах олиготрофных осоково-сфагновых мочажин и составили 8 т/га. На мерзлых буграх запасы фитомассы достигли максимальной величины, на кочках - 24 т/га за счет вклада лишайников, запасы которых в два раза больше, чем в межкочьях. Вечнозеленые вересковые кустарнички, занимающие верхние части кочек, создают внутри куста ветровую тень, которая благоприятствует росту лишайников. Большая часть подземной живой массы (60-80%) на повышенных элементах рельефа была сформирована крупными и мелкими корнями кустарничков и их погребенными стволиками.
Запасы фотосинтезирующих частей мхов и лишайников достигли значительной величины и колебались в пределах от 3,8 до 4,0 т/га, что составило около 25% от общих запасов фитомассы (Рис. 4б). Запас подземных органов трав и кустарничков составил 60–70% от общего запаса растительного вещества. В мочажинах структура фитомассы изменилась. Лидирующие позиции перешли к сфагновым мхам и подземными органами трав, их количество в среднем составило около 6 т/га (Titlyаnova et al., 1999).
Во всех исследованных экосистемах наибольшие запасы надземной фитомассы, не менее половины, приходились на мхи и лишайники. Наибольшими запасами фитомассы мхов отличались мочажины, а кочки бугров – наибольшими запасами фитомассы лишайников. С повышением трофности в мезотрофных мочажинах хасыреев запасы фитомассы мхов возросли до 2,1 т/га. На буграх приземный ярус сложен лишайниками, запасы фотосинтезирующей фитомассы которых составляли 4.0 т/га. В целом наибольшие запасы фитомассы оказались в экосистемах с относительно высокой трофностью – в мезотрофных хасыреях (18,5 т/га); в олиготрофной мочажине запасы снизились до 8,0 т/га.
Чистая первичная продукция экосистем плоскобугристого болота колебалась в пределах от 3,0 до 4,5 т/га в год в зависимости от видового состава растительного сообщества и топографии. В среднем она составила около 4,0 т/га в год при запасе фитомассы – 14,8 т/га, в хасыреях продукция достигла максимальной величины – 8,5 т/га в год, при запасах фитомассы – 18,0 т/га. На плоских буграх отношение надземной продукции к подземной, где доминировали корни кустарничков, получилось 1:4; в мочажинах, где преобладали подземные органы осок и пушиц, отношение увеличилось и составило 1:6. Наибольший вклад в продукцию внесли подземные органы во всех исследуемых экосистемах (рис. 5).
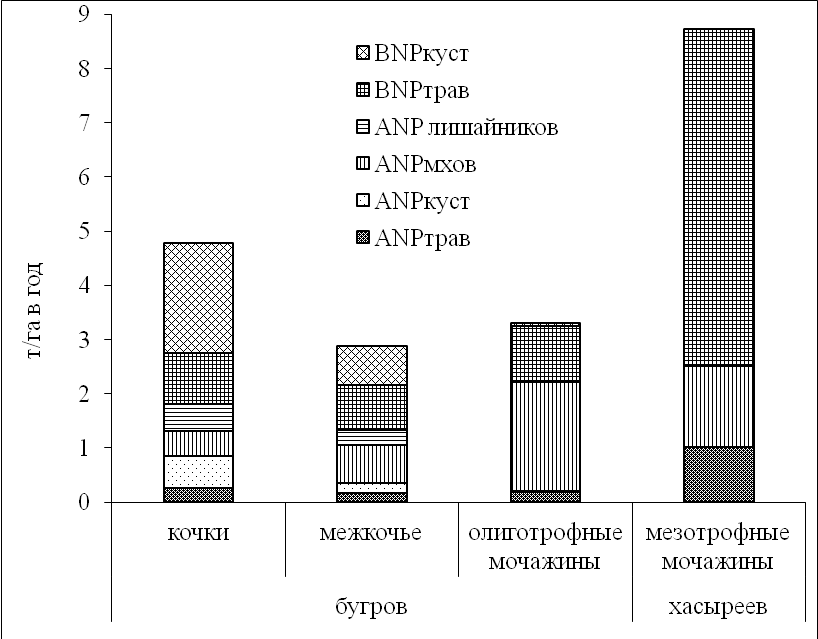
Рисунок 5. Чистая первичная продукция различных фракций растительного вещества на разных элементах рельефа плоскобугристого болота
Вклад корней трав составил в олиготрофных мочажинах 1,0 т/га в год, в хасыреях он возрос до 6,2 т/га в год, на буграх - 0,8-0,9 т/га в год. Вклад корней кустарничков на буграх варьировал от 0,7 до 2,0 т/га в год, в олиготрофных мочажинах составил 0,06 т/га в год. Вклад мхов и лишайников в продукцию экосистем менялся в пределах 1,0 - 2,0 т/га в год. Минимальный вклад в общую продукцию на повышенных и пониженных элементах рельефа внесли фотосинтезирующие части трав и кустарничков - 0,2 – 1,0 т/га в год (Косых, 2005).
Доля годичной продукции от общих запасов фитомассы во всех сообществах колебалась в пределах от 20 до 47%, и в целом подчинялась той же закономерности, что и распределение запасов фитомассы. Структура годичной продукции в изученных сообществах имела существенные различия, связанные с жизненной формой растений. Продукция трав и мхов во всех исследованных экосистемах составила меньше половины от общих запасов их живой фитомассы. Продукция кустарничков значительно меньше своих запасов из-за большого вклада многолетних побегов. Во всех сообществах продукция подземных органов растений оказалась выше продукции надземных.
Обсуждение
Продукционный процесс значительно полнее изучен нами в болотах средней и южной тайги (Косых и др., 2010; Kosykh et al., 2008). Наиболее продуктивными в средней тайге являлись мезотрофные болота (8,90 т/га в год). В этих экосистемах в формирование общей продукции равную долю (40%) внесли подземные органы трав и мхи. Надземная продукция трав и кустарничков не превышала 20%. В экосистемах рямов и олиготрофных мочажин доминантами продукционного процесса выступали сфагновые мхи. Их прирост преобладал в общей продукции экосистем. Самая низкая продукция фитомассы наблюдалась в олиготрофных мочажинах. В экосистемах гряд на верховом болоте в результате выноса питательных веществ поверхностным стоком сформировались бедные экосистемы, продукция которых не превышает 5,60 т/га в год.
Продукционный процесс на болотах Канады изучали Ридер, Стюарт (Reader, Stewart, 1972) и Мор (Moore et al., 2002), на болотах Швеции - Валлен (Wallen, 1992) и на болотах Финляндии Меррей (Murray et al., 1993). По данным этих авторов чистая первичная продукция на изучаемых ими болотах варьировала в пределах от 3,5 до 19,4 т/га в год и имела тенденцию к повышению с севера на юг. В Финляндии Джелмер (Nijp et al., 2014) определил запасы фитомассы на болотах Скандии, ее величина колебалась от 1,50 до 3,4 т/га. Сааринен (Saarinen, 1996) показал, что продукция корней Carex в травяных болотах составила 8,70 т/га в год, из которых на надземную продукцию приходилось 25% и на подземную – 75%. В болотах Южной Швеции продукция составляла 8,00 т/га в год, где на долю корней приходилось более 70 % (Wallen, 1992). Для болот северной тайги Западной Сибири доля подземных органов в продукции составляла в среднем 60% от общей продукции. Для Канады (Beilman et al., 2001) приводились данные общей продукции для северных болот: от 3,70 (плоскобугристые болота) до 4,50 т/га в год (олиготрофные болота). Полученные нами данные продукции для плоскобугристых болот северной тайги Западной Сибири не противоречат количественным характеристикам первичной продукции, полученным для тех же болот Канады, Швеции и Финляндии.
Заключение
В условиях современной нагрузки на переувлажненные ландшафты северных территорий возникают негативные последствия, вызывающие деградацию мерзлоты и связанные с ней процессы смены функционирования болотных комплексов.
При проведении работ по определению запасов и продукции надземного и подземного растительного вещества болотных экосистем рассматривали распределение его в активном слое почвы (ежегодно оттаиваемый почвенный слой).
Исследования растительности и растительного вещества плоскобугристых болот показали, что средние значения запасов растительного вещества как в надземной, так и в подземной сфере сохранялись в течении ряда лет на одном уровне и соответствуют средним значениям продукции, полученным для болот Канады, Швеции и Финляндии.
Распределение запасов фитомассы и годичной продукции в изучаемых болотных экосистемах северной тайги зависило от условий минерального питания почвы и обводненности корнеобитаемого слоя. Доминирующая роль мхов в продукции экосистемы сохранялась только в олиготрофных мочажинах, на буграх преобладали лишайники, видовое разнообразие которых превосходило остальные группы растений. Максимальный вклад подземной продукции наблюдался в мезотрофных мочажинах и составлял 70 % от общей продукции; этот вклад определяли, в основном, подземные органы крупных осок и пушиц. Общая первичная продукция на буграх составляла 4,1 т/га в год, что в два раза ниже, чем на грядах болотных комплексов средней тайги.
При равных условиях в мочажинах и хасыреях запасы фитомассы и годичная продукция растут с увеличением трофности почвы, а в экосистемах бугров уменьшаются при наличии мерзлого слоя почвы. Анализ продуктивности на разных болотных почвах северной тайги позволил сделать вывод, что почвенные условия оказывают существенное влияние на интенсивность продукционного процесса, обусловливая в целом ее невысокий уровень. При среднем общем запасе биомассы, которая составляла 170-220 т/га, годичный прирост в надземной сфере изменялся в пределах 1,3–2,5 т/га в год, а в подземной 1-6 т/га в год.
Благодарности
Работа выполнена при финансовой поддержке проекта РФФИ 18-44-700005 (работы по отбору проб и на экспериментальных площадках болотных комплексов для определению запасов и продукции растительного вещества); междисциплинарного интеграционного проекта СО РАН №45 (выбор экспериментальных площадок в экотонах лес-болото, отбор проб почв и растительного вещества для аналитических работ), а также проекта РФФИ 18-05-60264 Арктика.
Литература
- Андреев В.Н. Прирост кормовых лишайников и приемы его регулирования // Труды Бот. ин-та АН СССР. Сер. 3. Геоботаника. Вып. IX. 1954. С. 11-74
- Андреяшкина Н.И., Андреяшкина Ю.Г. Разногодичная изменчивость продукции надземной биомассы тундровых сообществ Зауралья // Структура и функционирование биогеоценозов Приобского Севера. Свердловск: Изд-во УНЦ АН СССР, 1981. С.12-19
- Базилевич Н.И., Титлянова А.А. Биотический круговорот на пяти континентах: азот и зольные элементы в природных наземных экосистемах. Новосибирск: Из-во СО РАН, 2008. 376с.
- Брыксина Н.А., Полищук Ю.М. Анализ изменения численности термокарстовых озер в зоне многолетней мерзлоты Западной Сибири на основе космических снимков // Криосфера Земли. 2015. T. 19. № 2. С. 114–120. Полный текст
- Василевская В.Д., Иванов В.В., Богатырев Л.Г. Почвы Севера Западной Сибири. М. Изд-во МГУ, 1986. 226 с.
- ГОСТ 10650-2013. Торф. Методы определения степени разложения. М.: Издательство Стандартинформ, 2014. 13 с. Полный текст
- Классификация и диагностика почв России / Авторы и составители: Л.Л. Шишов, В.Д. Тонконогов, И.И. Лебедева, М.И. Герасимова. Смоленск: Ойкумена, 2004. 342 с.
- Косых Н.П. Чистая первичная продукция болот Севера Западной Сибири // Биология и биосфера: Сборник материалов Четвертой Научной Школы (12-15 сентября 2005 г.) / Инишева Л.И. (отв. ред.). Томск: Изд-во ЦНТИ, 2005. С. 228-231.
- Косых Н.П., Вишнякова Е.К., Коронатова Н.Г. и др. Продукционно-деструкционные процессы в олиготрофных мочажинах средней тайги в ОТС-эксперименте // Углеродный баланс болот Западной Сибири в контексте изменения климата: Материалы Международной конференции (Ханты-Мансийск, 19-29 июня 2017). Томск: Изд-во ТГУ, 2017. С.77-7
- Косых Н.П., Миронычева-Токарева Н.П., Вишнякова Е.К. Продукционно-деструкционные процессы в болотных почвах // Отражение био-, гео-, антропосферных взаимодействий в почвах и почвенном покрове: Материалы IV Всероссийской научной конференции с международным участием. (Томск, 1-5 сент. 2010 г.). T. II: Томск: Изд-вл ТГУ, 20 С.105-108.
- Мельцер Л.И. Тундровая растительность // Растительный покров Западно-Сибирской равнины. Новосибирск: Наука, 1985. С. 41-54.
- Москаленко Н.Г. Антропогенная динамика растительности равнин криолитозоны России. Новосибирск: Наука, 1999. 280 с.
- Павлов А. В. Мониторинг криолитозоны. Новосибирск: Академическое изд-во «Гео», 2008. 299 с.
- Смоленцев Б.А. Структура почвенного покрова Сибирских увалов. Новосибирск: Изд-во СО РАН, 2002. 118с.
- Тишков А.А. Географические особенности первичных и вторичных сукцессий // Общие проблемы биогеоценологии: Тезисы II Всесоюзного совещания (Москва, 11-13 ноября 1986 г.). М.: НЦБИ АН СССР в Пущине, 1986. С.61-63.
- Тыртиков А.П. Динамика растительного покрова северной тайги Западной Сибири и вечная мерзлота // Природные условия Западной Сибири. М.: Изд-во МГУ, 1972. Вып.2. С. 100-114.
- Beilman D.W., Yu Z., Vitt D.H. Vodelling regional variability in peat accumulation response to climate in continental Western Canada // Торфяники Западной Сибири и цикл углерода: прошлое и настоящее: Тезисы Международного Симпозиума (Ноябрьск, 18-22 августа 2011). Васильев В.С., Tитлянова А.А., Величко А.А. (отв.ред.). Томск: Изд-во науч.-технич. литературы, 2001. С.15-18.
- Kosykh N.P., Koronatova N.G., Naumova N.B., Titlyanova A.A. Above- and below-ground phytomass and net primary production in boreal mire ecosystems // Wetlands ecology and management. 2008. Vol.16. №2. P. 139-153. CrossRef
- Moore T.R. Bubier J.L., Frolking S.E. et al. Plant biomass, production, CO2 exchange in an ombrotrphic bog// J.Ecol. 2002. V. 90. P.25
- Murray K.J., Tenhunen J.D., Nowak R.S. Photoinhibition as a control on photosynthtsis and production of Sphagnum mosses // Oecologia. 1993. Vol. 96. №2. P. 200-207. Crossref
- Nijp J.J., Limpens J., Metselaar K. et al. Can frequent precipitation moderate the impact of drought on peatmos uptake in northern peatlands? // New Phytol. 2014. Vol. 203., №1. P. 70-80. CrossRef
- Reader R.J., Stewart J.M. The relationship between net primary production and accumulation for a peatland in Southeastern Manitoba // Ecology. 1972. Vol.53. №6. P.1024-1037. CrossRef
- Saarinen T. Vascular plants as input of carbon in boreal sedge fens: control of production and partitioning of biomass. Helsinki, 1999. 66 p.
- Titlyаnova A.A., Romanova I.P., Kosykh N.P., Mironycheva-Tokareva N.P. Pattern and process in above-ground and below-ground components of grassland ecosystems // Journal of vegetation Science. 1999. Vol.10. Issue 3. P.307-320. CrossRef
- Wallen B. Methods for studying below-ground production in mire ecosystems // Suo. 1992. Vol. 43. №4-5. Р. 155-162.
- IUSS Working Group WRB, World Reference Base for Soil Resources 2006. A framework for international classification. № 103. FAO, Rome, 2006. 144 р. Полный текст
Сведения об авторах:
Косых Наталья Павловна – кандидат биологических наук, старший научный сотрудник лаборатории биогеоценологии ФГБУН Института почвоведения и агрохимии СО РАН (г. Новосибирск, Россия); kosykh@issa-siberia.ru
Миронычева-Токарева Нина Петровна – к.б.н., с.н.с. кафедры почвоведения и оценки земельных ресурсов Академии биологии и биотехнологии им. Ивановского Д.И при Южном Федеральном университете (г.Ростов-на-Дону, Россия); mirtok@issa-siberia.ru
Михайлова Елена Владимировна – кандидат географических наук, старший преподаватель кафедры экологии и природопользования Сибирского государственного университета геосистем и технологий; научный сотрудник лаборатории биогеоценологии ФГБУН Института почвоведения и агрохимии СО РАН, (г. Новосибирск, Россия); milek123@mail.ru
Колесниченко Лариса Геннадьевна – кандидат биологических наук, старший научный сотрудник лаборатории биоразнообразия и экологии Научно-исследовательского института биологии и биофизики Томского государственного университета (г. Томск, Россия); klg77777@mail.ru
Авторы прочитали и одобрили окончательный вариант рукописи
 Статья доступна по лицензии Creative Commons Attribution 4.0 License
Статья доступна по лицензии Creative Commons Attribution 4.0 License
VEGETATION AND PLANT MATERIAL OF THE FLAT PALSA PEATLANDS
N.P. Kosykh1 , N.P. Mironycheva-Tokareva1, E.V. Mikhailova1, 2, L.G. Kolesnichenko3
Address:1Institute of Soil Science and Agrochemistry of Siberian Branch of the Russian Academy of Sciences, Novosibirsk, Russia
2 Siberian State University of Geosystems and Technologies, Novosibirsk, Russia
3Tomsk State University, Tomsk, Russia
The aim of the study was to reveal peculiarities and regularities in plant material stock and production as dependent on the plant cover composition, soil characteristics and microrelief of the flat palsa mires in the northern taiga zone in West Siberia. The hollow-lake complexes on permafrost peatlands were chosen as the study sites. The work was carried out in 2013-2017. Plant material structure with differentiation between living and dead belowground organs was investigated and net primary production evaluated. Net primary production was found to range 3.0-4.5 t/ha per year depending on plant species composition of the phytocenosis studied. The production averaged 4.0 t/ha per year with phytomass stock averaging 14.8 t/ha, whereas in dried lake sites the net primary production reached 8.5 t/ha per year with phytomass stock estimated as 18.0 t/ha. On flat palsa sites the ratio of the aboveground shrub production to their belowground production was 1:4, while in hollow sites the belowground organs of sedges and cotton grasses accounted for most of the production, with the ratio of the above- to the belowground production estimated as 1:6. Belowground production share in the total production was found to be maximal (70%) in mesotrophic hollows mainly due to the major sedges and cotton grasses. The total primary production was estimated as 4.1 t/ha per year on flat palsa sites, 3.0 t/ha per year in oligotrophic hollows and reached a maximum of 8.5 t/ha per year in dried lake sites. Thus the average plant material stocks, both above- and belowground, seemed to be rather constant from year to year, and their estimates agreed well with those reported for mires in Canada, Sweden and Finland. Phytomass stocks and production in flat palsa mires in the northern taiga of the West Siberia were shown to depend on soil mineral nutrition and water availability in the root-inhabited layer. Only in oligotrophic hollows mosses were found to dominate in production, while the latter on the palsa sites was dominated by lichens, displaying great species diversity, significantly exceeding that of other plants. The other factors being equal, in hollow sites phytomass stocks and yearly production increased with increased nutrient availability, being decreased on palsa sites due to the permafrost layer. Analysis of phytomass production in northern taiga mires allows concluding that soil characteristics play important role in phytomass production, which was found to be rather low overall. Phytomass stock and production on palsa sites are limited by shallow thawing depths, which effect is exacerbated by drying and warming of the upper soil layer in summer, altogether resulting in unfavourble environment for root growth and development.
Key words: peatlands; permafrost; biological production; plant material; phytomass stock; mortmass stock; net primary production
How to cite: Kosykh N.P., Mironycheva-Tokareva N.P., Mikhailova E.V., Kolesnichenko L.G. Vegetation and plant material of the flat palsa peatlands // The Journal of Soils and Environment. 2019. 2(1). e55. doi: 10.31251/pos.v2i1.55 (in Russian with English abstract).
References
- Andreev V.N. Growth of forage lichens and methods of its regulation In book: Proc. of Botanic Institute of the USSR AS. Ser. 3 Geobotany. Issue IX, 1954, p.11-74. (in Russian)
- Andreiashkina N.I., Andreiashkina Yu.G. Multi-year variability of above-ground biomass production in tundra communities of Trans-Urals In book: Structure and functioning of biogeocenoses of the Priobsky North. Sverdlovsk: Publ. House of the UNC of the USSR AS, 1981, p.12-19. (in Russia)
- Bazilevich N.I., Titlyanova A.A. Biotic turnover on five continents: element exchange processes in terrestrial natural ecosystems. Novosibirsk, Publ. SB RAS, 2008, 376 p. (in Russian)
- Bryksina N.A., Polishchuk Yu.M. Analysis of the change in the number of thermokarst lakes in the permafrost zone of Western Siberia based on satellite images, Earth’s Cryosphere, 2015, Vol. 19, No 2, p. 114–120.(in Russian) Full text
- Vasilevskaya V.D., Ivanov V.V., Bogatyrev L.G. Soils of the North of Western Siberia. Moscow, Publ. House of MSU, 1986 ,226 с. (in Russain)
- GOST 10650-2013. Peat Methods for determining the degree of decomposition. Moscow: Standard Publishing House, 13 p. (in Russian) Full text
- Soil classification and diagnostic of Russia / Authors and compilers: L.L. Shishov, V.D. Tonkonogov, I.I. Lebedeva, M.I. Gerasimov. Smolensk: Oykumena Publ., 2004. 342 p. (in Russian)
- Kosykh N.P. Net Primary Production marshes of the North of Western Siberia In book: Biology and the Biosphere: Proc. of the 4th Sci. School (Tomsk, 12-15 September 2005). Inisheva L.I. (Ed.). Tomsk: Publishing House of CSTI, 2005, p. 228-231. (in Russian)
- Kosykh N.P., Vishnyakova E.K., Koronatova N.G. et al. Production-destruction processes in oligotrophic hollows in the middle taiga in the OTS experiment In book: Carbon Balance of Marshes in Western Siberia in the Context of Climate Change: Proc. of the Inter. Conf. (Khanty-Mansiysk, June 19-29, 2017). Tomsk: Publ. House of TSU, 2017, p.77-7 (in Russia)
- Kosykh N.P., Mironycheva-Tokareva N.P., Vishnyakova E.K. Production and destruction processes in bog soils In book: Reflection of Bio-, Geo-, Anthropospheric interactions in soils and soil cover: Proc. of the IV Rus. Sci. Conf. with Inter. participation. (Tomsk, September 1-5, 2010). Vol. II, Tomsk, 20 p. 105-108. (in Russian)
- Melzer L.I. Tundra vegetation In book: Vegetation cover of the West Siberian Plain, Novosibirsk: Science Publ., 1985, p. 41-54. (in Russian)
- Moskalenko N.G. Anthropogenic dynamics of vegetation of plains in russian cryolithozone. Novosibirsk: Science Publ., 1999, 280 p. (in Russian)
- Pavlov A.V. Monitoring of the cryolithozone. Novosibirsk: Academic Publ. House "Geo", 2008, 299 p. (in Russain)
- Smolentsev B.A. The structure of the soil cover of the Siberian ridges. Novosibirsk: Publ. House of the SB RAS, 2002, 118 p. (in Russian)
- Tishkov A.A. Geographical features of primary and secondary successions In book: General problems of biogeocenology: Book abstracts of the II Soviet Union Conf. (Moscow, 11-13 November 1986). Moscow.: NCBI AS of the USSR in Pushchin, 1986, p. 61-63. (in Russian)
- Tyrtikov A.P. Dynamics of the vegetation cover of the northern taiga of Western Siberia and the permafrost in book: Natural conditions of Western Siberia. Moscow.: Publ. House of MSU, 1972, Issue 2, p. 100-114. (In Russian)
- Beilman D.W., Yu Z., Vitt D.H. Vodelling regional variability in peat accumulation response to climate in continental Western Canada In book: West Siberian peatlands and Carbon cycle: past and present: Book abstracts of the International Field Simposium (Noyabrsk, 18-20 August, 2001). Vasiliev V.S., Titlyanova A.A., Velichko A.A. (Eds.). Tomsk: Publ. House of Sci. and Tech. Literature, 2001, p.15-18.
- Kosykh N.P., Koronatova N.G., Naumova N.B., Titlyanova A.A. Above- and below-ground phytomass and net primary production in boreal mire ecosystems, Wetlands ecology and management, 2008, Vol.16, No2, p. 139-153. CrossRef
- Moore T.R. Bubier J.L., Frolking S.E. et al. Plant biomass, production, CO2 exchange in an ombrotrphic bog// J.Ecol. 2002. V. 90. P.25
- Murray K.J., Tenhunen J.D., Nowak R.S. Photoinhibition as a control on photosynthtsis and production of Sphagnum mosses, Oecologia, 1993, Vol. 96. No2, p. 200-207. CrossRef
- Nijp J.J., Limpens J., Metselaar K. et al. Con frequent precipitation moderate the impact of drought on peatmos uptake in northern peatlands?, New Phytol, 2014, Vol. 203, №1, p. 70-80. CrossRef
- Reader R.J., Stewart J.M. The relationship between net primary production and accumulation for a peatland in Southeastern Manitoba, Ecology, 1972, Vol.53, No6, p.1024-1037. CrossRef
- Saarinen T. Vascular plants as input of carbon in boreal sedge fens: control of production and partitioning of biomass. Helsinki, 1999. 66 p.
- Titlyаnova A.A., Romanova I.P., Kosykh N.P., Mironycheva-Tokareva N.P. Pattern and process in above-ground and below-ground components of grassland ecosystems, Journal of vegetation Science, 1999, Vol.10, Issue 3, p.307-320. CrossRef
- Wallen B. Methods for studying below-ground production in mire ecosystems, Suo, 1992, Vol. 43. Issue 4-5, p. 155-162.
- IUSS Working Group WRB, World Reference Base for Soil Resources 2006. A framework for international classification. № 103. FAO, Rome, 2006. 144 р. Full text
Received 10 December 2018; accepted 26 April 2019; published 27 April 2019
About the authors:
Kosykh Natalia P. – Candidate of Biological Sciences, Senior Researcher of the Laboratory of Biogeocenology Institute of Soil Science and Agrochemistry, Siberian Branch of the Russian Academy of Sciences (Novosibirsk, Russia); kosykh@issa-siberia.ru
Mironycheva-Tokareva Nina P. – Candidate of Biological Sciences, Associate Professor, Head of the Laboratory of Biogeocenology, Institute of Soil Science and Agrochemistry, Siberian Branch of the Russian Academy of Sciences (Novosibirsk, Russia); mirtok@issa-siberia.ru
Mikhailova Elena V. – Ph.D. (Geography), Senior Lecturer, Department of Ecology and Environmental Management, Siberian State University of Geosystems and Technologies; Research Associate in the Laboratory of Biogeocenology, Institute of Soil Science and Agrochemistry, Siberian Branch of the Russian Academy of Sciences, (Novosibirsk, Russia); milek123@mail.ru
Kolesnichenko Larisa G. – Candidate of Biological Sciences, Senior Researcher of the Laboratory of Biodiversity and Ecology of the Research Institute of Biology and Biophysics of Tomsk State University (Tomsk, Russia); klg77777@mail.ru
The authors read and approved the final manuscript
The article is available under Creative Commons Attribution 4.0 License